
1972年Burnstock首次提出体内存在有嘌呤受体这一设想,1978年“嘌呤”受体(“Purinoceptor”)被正式命名,并被进一步区分为P1(腺苷)和P2(ATP)受体。P1受体再分为A1、A2、A3等亚型,而P2受体则分为P2X和P2Y两大类。1991年陆续从不同种属动物克隆出数种具有功能性的P2Y受体;P2X受体自1994年后也克隆出P2X1-7种亚基。以往的其它分类名称,如P2D,P2T,P2U等均已归入P2Y受体,而P2Z则属配体门控离子通道受体。2011年,美国科学家首次发现,老鼠身体内产生的一种分子——腺嘌呤核苷受体能对大分子进入大脑进行控制,当腺嘌呤核苷受体在组成血脑屏障的细胞上被激活时,就会建立起一个进入血脑屏障的通道。
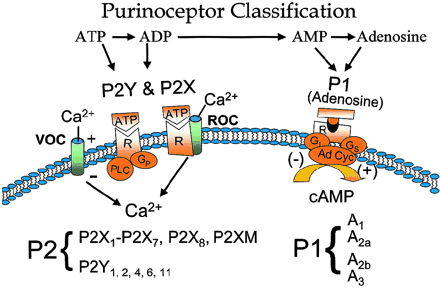
P1 and P2 receptor classification and the signaling mechanisms thought to be involved. PLC, phospholipase C; GP, G proteins; R, receptor; VOC, voltage-operated calcium channel; ROC, receptor-operated calcium channel; GI, inhibitory G proteins; GS, stimulatory G proteins; Ad Cyc, adenylate cyclase.在实验中,研究人员成功地将葡萄聚糖和抗体一样大小的大分子运送至大脑中,试图厘清它们能让大分子到达何处以及这种方法是否对分子的大小有要求。他们也成功地让一个β淀粉样肽抗体穿过转基因老鼠的血脑屏障,并观察到它依附于导致老鼠罹患阿尔茨海默病的淀粉状蛋白斑上。在老鼠体内,还有很多已知的对抗剂(专门阻止信号传递的药物或者蛋白)可作腺嘌呤核苷受体。
P2嘌呤受体最初被分为P2X和P2Y两种亚型,其后扩展到P2T、P2Z和P2D亚型。90年代发现,一些组织标本对UTP、ATP和ATPγS反应良好,而对a,β-MeATP和2-MeSATP不敏感,此类受体被称为“P2U受体”。晚近又证明,存在一种对UTP敏感而对ATP不敏感的“嘧啶受体”。据此IUPHAR(International union of pharmacology)规定,任何被核苷酸激活的离子通道型和G蛋白偶联型受体的亚型均分别命名为P2Xn和P2Yn受体。随着分子生物学技术的发展以及上述受体的克隆和表达,此分类系统得到了有力的支持。 P1和P2Y受体是G蛋白偶联受体,P2X受体是配体门控的离子通道;嘌呤受体在神经系统和外周组织广泛存在,执行多种生理功能:疼痛的产生和调制,神经系统的发育和再生,内皮细胞介导的血管舒张,调节腺体分泌,血小板聚集,调节神经元、AS及小胶质细胞功能,免疫调节,炎症反应,调节记忆功能,调节递质释放等;嘌呤受体与多种神经递质存在共释放(包括:Glu、去甲肾上腺素、乙酰胆碱、GABA、5-HT等)和相互作用。在CNS,嘌呤受体表达或功能异常会导致神经元和胶质细胞损害,而CNS损伤(TBI和脊髓损伤)和神经系统退行性改变也会导致胞外ATP释放增加以及嘌呤受体表达和功能改变,进一步加重CNS损伤。

ATP及其类似物经由激活不同的嘌呤受体影响免疫细胞对炎症的反应,其中P2X7受体有强的促前炎症因子释放作用,比如促进巨噬细胞样细胞释放IL-1β,此作用被认为是加重继发性脑损伤,促进细胞死亡的因素之一。Neary, J. T.等报道,TBI后ATP快速释放,通过作用于P2受体,促进AS的存活和再生,在TBI后的胶质化中起重要作用。在海马缺血性实验中,阻断P2X受体对海马CA1区神经元有保护作用。Shirakawa, T.等报道,海马CA1区P2X1受体参与了神经毒性物质TMT诱导的锥体细胞死亡。Franke,H.等的研究显示,在神经系统炎症的发生中,ATP与Glu、GABA有协同作用;其中P2X和P2Y受体分别介导突触的快反应和慢反应。Wagner, A. K.等报道,严重TBI后海马嘌呤A1受体的基因改变参与了PTE的形成。本课题组前期研究发现,在苦味毒诱发的癫痫实验中,中药大黄的主要成分大黄素通过作用于海马CA1区的A1受体,抑制Glu的释放和EPSP。
Koles, L.等研究报道,神经元和胶质细胞间(包括AS和小胶质细胞)的相互作用介质主要是ATP和Glu,且主要通过作用于P2X7和P2Y受体而实现。P2X7受体是P2X受体家族中非常独特的一个亚类,特点是:①活化时所需的ATP浓度较高(>100μM);②P2X7离子通道激活后可以形成一个大的非选择性细胞孔,可通过分子量较大(900Da)的物质。Sperlagh, B.等在培养的海马脑片中发现,激活CA1和CA3区兴奋性神经末梢的P2X7受体能调节Glu和GABA的释放。同时在P2X7R(-/-)大鼠海马,ATP不能激发Glu和GABA的释放,说明海马ATP引起的Glu和GABA的释放由P2X7受体介导。进一步研究表明,ATP作用于海马P2X7受体引起的Glu释放由p38MAPK介导。在CA1锥体细胞,P2X7受体介导非Ca2+依赖性的Glu释放,在病理状态下此种作用被增强导致CNS损伤。Labrousse, V. F.等报道,海马P2X7受体可调控IL-1β的合成,参与调节神经系统炎症反应。Kim, J. E.等报道,P2X7受体通过TNF-α途径改善癫痫持续状态下的海马CA3区神经元的损伤程度。Jacques-Silva, M. C.等发现,在培养的皮层AS,ATP及其类似物BzATP促进AS蛋白激酶AKt的磷酸化的作用由P2X7受体介导。在海马LPS诱导的炎症实验中,P2X7受体介导了小胶质细胞IL-1β的释放,增加组织对兴奋性损伤的易感性,促进小胶质细胞死亡。Oliveira, J. F.等也报道,CNS损伤时ATP释放增加,作用于AS的P2X7受体,参与坏死/凋亡或增殖反应。




